Фосфор и кальций
Фосфор и кальций настолько тесно связаны между собой в обмене веществ, что их необходимо рассмотреть вместе. Значение фосфора для организма животного достаточно известно. Он является главным участником во всех жизненно важных процессах обмена веществ и поэтому встречается в каждом биологическом материале. Фосфор является структурным элементом костной и зубной тканей: у молодняка здесь содержится 83%, а у взрослого крупного рогатого окота 87% всего количества фосфора, находящегося в организме.
В форме щелочных и кислых фосфатов фосфор включен в буферные системы, входит в состав соединений, необходимых для обмена энергии, как, например, аденозинтрифосфорная кислота (АТФ) или фосфокреатин, и является структурным элементом нуклеиновых кислот, роль которых в обмене веществ в клетке и передаче наследственной информации в последние годы проявляется все более отчетливо. Во время охоты и течки в промежуточном мозге, гипофизе и яичниках - органах, которые управляют половой функцией,- обмен фосфатов повышается. В сперме тоже много фосфорсодержащих [259] белков. К сожалению, на такое широкое значение фосфора в питании домашних животных, особенно в питании крупного рогатого окота, обращают слишком мало внимания.
Около 99% содержащегося в организме кальция сосредоточено в скелете и зубах, остальная часть - преимущественно в жидкостях организма. Так, концентрация кальция в плазме крови составляет около 9-12 мг %, половина его связана с белками. Неорганическое вещество костей состоит преимущественно из трехзамещенного фосфата кальция, отношение Са : Р составляет примерно 2 : 1.
Наряду с этим кальций выполняет важную функцию в возбуждении нервной системы и мускулатуры. При окислительном декарбоксилировании он активирует некоторые ферменты и липазы, изменяет проницаемость клеточных мембран. Присутствие кальция необходимо для превращения протромбина в тромбин, в чем выражается его влияние на свертываемость крови.
Фосфор и кальций всасываются преимущественно в тонком отделе кишечника и лишь в ограниченном размере в желудке. В кормах фосфор присутствует всегда в форме фосфатов, поэтому более правильно было бы говорить о резорбции фосфатов. Уровень всасывания обоих элементов зависит от нескольких факторов. Так, переход фосфатов из слизистой оболочки в серозную происходит в присутствии кальция и кислорода, в средах этот процесс стимулируется калием и угнетается цианидом.
Хотя витамин D ускоряет переход фосфора из слизистой оболочки в серозную, но и здесь необходимо присутствие кальция. Механизм действия витамина D полностью еще не ясен, но известна его регулирующая роль в процессе всасывания кальция и фосфатов из кишечника. При излишке кальция в рационе он стимулирует всасывание фосфатов и тормозит всасывание кальция. У поросят он, наоборот, часто стимулирует резорбцию кальция, так как в рационах фосфора нередко больше, чем кальция.
Из неорганических растворимых соединений фосфора всасывается, как правило, больше, чем из органических. "Истинная" переваримость фосфора из кормов или минеральных смесей достигает примерно 50-60%. Хорошо используется ортофоофат, всасывание же фосфора из конденсированных фосфатов (пиро-, поли- и метафосфатов) зависит от вида животных и от того, какой элемент выступает в фосфате в роли катиона. Если это кальций, то использование фосфора будет минимально или неудовлетворительно. Уровень всасывания при достаточном обеспечении витамином D и нормальном здоровье определяется потребностью в нем животных, т. е. у молодых, беременных и лактирующих особей кальция резорбируется больше, чем у старых, небеременных и нелактирующих. На уровень всасывания кальция помимо количества белка оказывает влияние и его биологическая ценность. При отсутствии в корме азота кальций почти не всасывается. Всасыванию его способствует включение в рацион лактозы. У очень молодых животных неблагоприятное влияние на резорбцию кальция оказывают фитин и фитиновая кислота.
Долго продержавшееся предположение, что щавелевая кислота, например из свеклы или свекольной ботвы, снижает интенсивность всасывания кальция из-за образования с ним труднорастворимого соединения оксалата, не подтвердилось, так как последнее расщепляется микрофлорой преджелудков и толстого отдела кишечника. Неодинаковая доступность кальция из отдельных кормов, установленная по меньшей мере для крупного рогатого скота, перекрывается отмеченными уже влияниями возраста, беременности и лактации.
Выделение фосфора из организма у растительноядных животных происходит главным образом с калом, а у всеядных еще и с мочой. Значительное выделение этого элемента происходит с молоком и слюной. Кальций тоже выделяется с калом, мочой, слюной, молоком и, кроме того, с потом.
Тейлер и Грин были первыми, кто высказал предположение о существовании заболеваний, связанных с недостатком фосфора. Они проследили связь между недостатком фосфора и заболеванием крупного рогатого скота на пастбищах Южной Африки, сопровождающимся явлениями атаксии, спастическими параличами и тяжелыми нарушениями воспроизводства.
Последующие сообщения о заболеваниях в связи с недостатком фосфора тоже поступили преимущественно из Австралии, Кении, Гвианы (французской). Сейчас в интернациональной литературе имеются многочисленные данные о том, что полное отсутствие фосфора в рационе нарушает развитие и здоровье животных. Тенденция к повышению уровня продуктивности, обусловленная экономическими соображениями, способствовала тому, что у крупного рогатого скота эта ситуация угрожающе обострилась. Выведение высокопродуктивного типа животных с более тонким телосложением несомненно уменьшило их способность адаптироваться к периодам временного недостатка или временно повышенной потребности в минеральных веществах в соответствии именно с этой высокой продуктивностью. Запасы кальция и главным образом фосфора в костяке стали более скудными, ежедневно увеличивается выделение их с молоком (на 1 кг молока примерно 1 г фосфора).
Симптомы фосфорной недостаточности у крупного рогатого окота обобщены по литературным данным [92] и приведены в таблице 9.
| Устанавливаются владельцем | Диагностируются ветеринарными специалистами |
|---|---|
| Худшая поедаемость кормов | Падение содержания жира в молоке |
| Резкий спад молочной продуктивности | Понижение уровень неорганического фосфора |
| Замедленный рост | Часто повышен уровень кальция в крови |
| Предрасположенность к маститам | Пониженная минерализация костей |
| Слабожизненные телята (нарушение пищеварения) | Шатание резцов |
| Послеродовой парез | Высокая степень деминерализации зубов и костей |
| Атаксии и параличи | Атрофия суставных хрящей |
| Кифоз или лордоз | Остеомаляция |
| Нервные симптомы | Рахит (у молодых животный) |
| Остеопороз | Тендовагиниты |
| Частичное облысение и депегментация | Анемия |
| Анафродизия вследствии пониженной функции яичников | (Гиперкератоз) |
| Нарушение зрения |
У свиней и крупного рогатого окота эти симптомы сходны. Особенно опасно это для растущих животных, так как, например, при содержании в корме только 0,1% фосфора развивается рахит. С другой стороны, для возникновения и течения такого рода заболеваний у крупного рогатого окота и свиней типично то, что клиническая картина с каждой последующей беременностью становится более резко выраженной.
Как уже подчеркивалось вначале, (вопросы обеспечения сельскохозяйственных животных кальцием и фосфором следовало бы рассматривать по существу в их взаимодействии друг с другом. К тому же симптомы дефицита кальция отчасти сходны с симптомами дефицита фосфора. Правда, чистые заболевания на почве недостатка кальция наблюдались до сих пор очень редко, чаще они вызываются избытком этого элемента. Высокие дозы кальция неблаготворно сказываются на потреблении норма. Особенно чувствительны к этому свиньи, в то время как жвачные переносят большие дозы кальция лучше всех прочих видов животных. Избыток кальция вызывает у свиней павакератоз, так как при этом нарушается распределение в организме цинка, а также магния, фосфора, железа, марганца и йода.
Хотя ни один из минеральных элементов не оказывает строго специфического влияния на половую сферу, фосфору на основании его выдающейся роли в промежуточном обмене веществ все же приписывают воздействие на процесс воспроизводства. Это положение, выдвинутое вначале Тейлером, было в деталях дополнено другими исследователями [18, 113, 114, 115, 118, 119, 214]. Наиболее глубоко изучили С. Хигнетт и П. Хигнетт влияние фосфора, кальция и витамина D на размножение. Самую низкую плодовитость они обнаружили в стадах с высоким потреблением кальция, но также и в тех, где был отмечен избыток фосфора в рационе, что позднее установил Конерманн [150]. При суточной даче 15-20 г Р2О5 (что соответствует примерно 6,6-8,6 г фосфора*) они устанавливали у телок анафродизию.
* (Фосфорный ангидрид (полупятиокись фосфора) используется в качестве эквивалента, показывающего наличие фосфора в кормах или удобрениях. В чистом виде как фосфорная подкормка не применяется.- Прим. перев.)
Неблагоприятное воздействие избытка фосфора на плодовитость Хенниг и Анке [103] обосновывают снижением обмена марганца, которое четко устанавливается анализом волосяного покрова животных при избыточном потреблении фосфора. Благотворное влияние на воспроизводство очень богатой фосфором костной муки, о чем писали Тейлер и в последнее время Грунер и Саитиаго [88], Хенниг с сотрудниками [106] не считают доказательством того, что фосфор необходим для процесса воспроизводства. Так как недостаток фосфора выражается еще в начальной стадии в уменьшении приема корма, то оплодотворяемость подопытных животных могла повыситься в результате возросшего потребления корма после добавок фосфора. Дальнейшее объяснение надо искать в присутствии в костной муке марганца. Вначале предполагалось, что недостаток фосфора только тогда приводит к бесплодию, когда уже проявились другие признаки, прежде всего сильное истощение. Однако, по мнению О'Моора [1950], выраженной фосфорной недостаточности предшествует так называемая субклиническая стадия, во В1ремя которой наблюдается только нарушение воспроизводства (субклинический афосфороз). При сравнении минерального статуса стад с нормальной и пониженной плодовитостью в рационах последних всегда обнаруживался недостаток фосфора и натрия. В стадах, хорошо обеспеченных минеральными (веществами, индекс осеменения составлял 1,3, а отход из-за бесплодия - 20%. Такова же и доля животных с нарушенным половым циклом. Те же показатели для стад, недостаточно обеспеченных минеральным питанием, составили 3,2 и 60%. Около 70% животных имели нарушенный половой цикл. На исходе зимы примерно у трети животных, пришедших в охоту, наблюдались запаздывание овуляции или атрезия фолликулов.
Хамблох [94] тоже установил в своих опытах, что в 20 стадах с нормальным воспроизводством потребность в фосфоре удовлетворялась на 108%, а в 20 других хозяйствах, где было нарушено воспроизводство, она удовлетворялась только на 79%.
Хидироглоу и др. [109] в явлениях бесплодия видят наряду с замедлением роста и типичные признаки острой фосфорной недостаточности. Появление метроррагий после эструса, которыми сопровождается бесплодие, Стефанов [239] объясняет недостатком фосфора в рационе. Яковач и др. [129] тоже рекомендуют искать причины нарушения воспроизводства крупного рогатого скота в остром дефиците фосфора. В Швейцарии, по наблюдениям Хаслера [98], число хозяйств с нарушением воспроизводства у животных возрастает особенно там, где при широком отношении Са : Р содержание фосфора в корме падает ниже 0,3%.
Штраух и Брюстнер [243] тоже установили в 57 хозяйствах прямую зависимость продолжительности межотельного периода и индекса осеменения от содержания фосфатов, в корме.
Исследования, проведенные на 123 стадах крупного рогатого скота Конерманном [150], отчетливо показали, что бесплодие, вызванное дефицитом фосфора, можно устранить улучшением обеспечения этим элементом. Правда, такая тенденция сохраняется лишь до наступления избытка 10 г в сутки. Всякое дальнейшее увеличение дозы фосфора сверх этого предела ведет к обратному действию, т. е. к снижению оплодотворяемости (рис. 8).
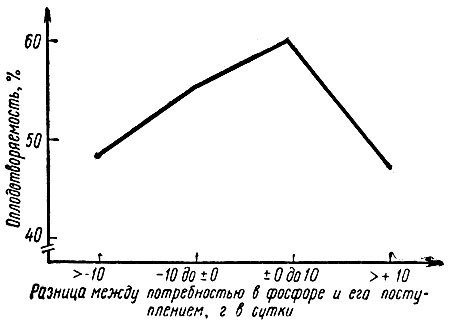
Рис. 8. Взаимосвязи между средним обеспечением животных фосфором и оплодотворяемостью по первому осеменению в 123 стадах крупного рогатого скота (по Конерманну, 1970)
Бече [13] пытается объяснить влияние фосфора на размножение, хотя бы у крупного рогатого окота, тем, что лактирующая корова в первую стадию лактации выделяет с молоком обычно больше минеральных веществ, чем может получить обратно с кормом. Следовательно, компенсация ее потребности в кальции и фосфоре будет зависеть в первую очередь от размеров и состава резервов минеральных веществ в этот очень критический для нее период, когда наряду с интенсивным образованием молока активно функционируют яичники.
Бели эти резервы скудны или уже исчерпаны, нарушается деятельность половых желез, так как молочная железа имеет преимущество в очередности обеспечения минеральными веществами. В этом причины нарушений полового цикла и оплодотворения, из-за которых удлиняется межотельный период и животное остается временно бесплодным. Период покоя половых функций сохраняется до тех пор, пока недостаток минеральных веществ не будет возмещен, т. е. пока не будут созданы условия для нормального протекания половых функций и тем самым для оплодотворения с последующей нидацией зиготы в матке. Другая ситуация наблюдается к концу лактации. Когда продукция молока снижается и минеральное равновесие организма вновь восстанавливается за счет потребления кормов с соответствующим содержанием кальция и фосфора, то необходимые количества этих элементов используются для пополнения израсходованных резервов и тем самым вновь создаются условия для нормальной половой деятельности.
Очень многие исследования, проведенные во всем мире, вновь и вновь указывают на то, что плодовитость сельскохозяйственных животных всегда выше там, где почвы богаче фосфорам и где вносится больше фосфатных удобрений на пастбища и пахотные земли хозяйства [188]. На 35 фермах Праворейнской возвышенности Мюллер [185] обнаружил, например, что в 1952 г. при содержании в почве 12 мг P2O5 на 100 г для одного оплодотворения требовалось в среднем 2-2,5 коитуса. Снижение содержания Р2О5 до 2 мг/100 г повысило число покрытий на одно оплодотворение до 3,5, однако отсутствие сведений о типе почвы не позволяет провести широкое сравнение с другими местностями. Не следует забывать и о том, что удобрение, например, томасшлаком одновременно обогащает почву марганцем, и здесь уже невозможно отграничить действие на воспроизводство фосфора и марганца [106].
Результаты своих дальнейших исследований Мюллер и Шюрманн [187] представили в виде таблицы (табл. 10).
| Содержание в 100 г почвы Р2О5 | 2 | 4 | 6 | 8 | 10 | 12 | 14 | 16 |
|---|---|---|---|---|---|---|---|---|
| Число осеменений на одно оплодотворение | 3,2 | 2,6 | 2,5 | 2,4 | 2,3 | 1,9 | 1,7 | 1,4 |
В серии подобных же исследований получены также данные о влиянии на воспроизводство крупного рогатого скота кальция и фосфора из минеральной подкормки (табл. 11).
| Вид подкормки | Число коров | Индекс бесплод |
|---|---|---|
| Только углекислая известь | 470 | 36 |
| Только соль-лизунец | 208 | 34 |
| Без минеральной подкормки | 3290 | 34 |
| Минеральная смесь: | ||
| с 8-12% Р2О5 | 4442 | 31 |
| с 16-18% Р2О5 | 348 | 27 |
* (Число дней сверх 365, составляющих период между отелами.)
В заключение упомянем об исследованиях Брюннера [34], который провел сравнение ряда дальнейших параметров с уровнем обеспеченности животных фосфором (табл. 12).
| Плодовитость хорошая | Плодовитость средняя | Плодовитость плохая | |
|---|---|---|---|
| Обследованно хозяйств | 14 | 26 | 17 |
| Количество исследованных коров | 339 | 813 | 453 |
| Индекс осеменения | 1,5 | 1,8 | 1,8 |
| Межотельный период, дни | 387 | 416 | 448 |
| Выбраковано всего, % | 7,1 | 13,8 | 18,3 |
| в том числе из-за бесплодия, % | 3,2 | 6,6 | 11,3 |
| Внесено с удобрениями Р2О5, кг/га | 37,6 | 32,7 | 22,8 |
| Содержание Р2О5 в 100 г почвы, мг | 6,4 | 5,5 | 4,2 |
| Содержание Р2О5 в сене, % | 0,45 | 0,42 | 0,38 |
Во всех такого рода и подобных им исследованиях дефицит фосфора большей частью сопровождался избытком кальция, нарушавшим благоприятное соотношение Са : Р, на что указывали также Хааб [91] и Кислинг [141]. Нередко, правда, отмечаются одновременно слишком высокий уровень калия и недостаток натрия в кормах. Поэтому мы подчеркиваем, что ссылка на недостаток фосфора как, на первичную причину бесплодия является недопустимым упрощением проблемы. Если учесть, что, например, у крупного рогатого скота с каждым литром молока выделяется 1,25 г кальция и 1 г фосфора, которые вначале должны быть усвоены организмом, то все равно оба эти элемента имеют особую ценность для молочных коров, несмотря на то, что в процессах воспроизводства кальций играет только второстепенную роль. Как показали американские опыты, плодовитость не нарушается от того, что корова на проятжении трех лет получала в рационе вместо рекомендованных 0,7% кальция только шестую часть этого количества - 0,12%. С другой стороны, некоторые формы бесплодия удавалось ликвидировать дачей углекислой извести (Русхофф и др., 1950). Шнейдевинд [221] и Кох (1934) наблюдали при гипокальциемии также гипоплазию яичников и удлинение течки. Ромбе (1962) на основании своих опытов пришел к заключению, что путем инъекции 40%-ного раствора глюконата кальция можно повысить оплодотворяемость коров, которые регулярно с трудом становились стельными.
В условиях сельскохозяйственной практики при содержании на обычных рационах из грубого корма, свекольной ботвы или бобовых можно скорее ожидать избытка, а не дефицита кальция. Так, коровы часто месяцами получают кальция в 5-10 раз больше необходимого им количества. Таким избытком кальция еще более обостряется фосфорная недостаточность, особенно при значительном использовании для кормления зерна и мельничных отходов.
В опытах на 34 коровах Конерманн [149] установил, что при избытке кальция сверх 110 г каждое последующее повышение его количества на 10 г ведет к удлинению межотельного периода и интервала от первого осеменения до оплодотворения на 16 дней (рис. 9). Наблюдавшееся в опытах других авторов [118] неблагоприятное воздействие больших количеств кальция на воспроизводство можно объяснить, по мнению Хеннига и др. [106], влиянием кальция на обмен некоторых микроэлементов, на что также указывал Альдерман [4].
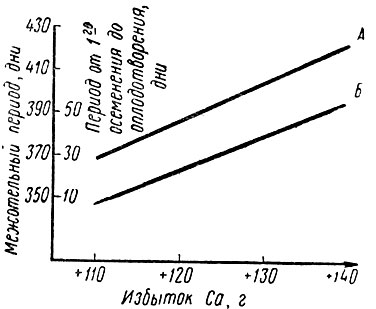
Рис. 9. Связь между плодовитостью и избыточным поступлением кальция (по Конерманну, 1970): А - межотельный период; Б - период от первого осеменения до оплодотворения
В последние годы в ГДР проведены исследования с целью выяснения прямых связей между обеспеченностью фосфором и плодовитостью женских особей жвачных животных. Так, Анке [6], Бугдоль [35], а также Хеиниг и др. [108] провели аналитические исследования кормовых растений с пастбищ на тех почвах Тюрингии, которые образовались в результате выветривания различных пород, и попытались найти связь между содержанием фосфора в растениях и плодовитостью коров по 1960 первичным осеменениям в зонах обслуживания станций искусственного осеменения Эрфурта и Йены. Позднее такие же исследования провели в Средней Саксонии Пелуг [204] и в округе Карл-Маркс-Штадт Лидлер [160]. В отличие от Мюллера [185] и Брюннера [34] ими было установлено, что содержание в почве фосфора в хозяйствах земли Северный Рейн-Вестфалия не коррелировало с результатами оплодотворения по первому осеменению. Самые лучшие результаты получены как в Тюрингии, так и в Саксонии в районах, расположенных на почвах с крайне неодинаковым содержанием фосфатов, а выдающиеся - на красном гнейсе с самой низкой оценкой по содержанию Р2О5. После экспериментальных исследований Хольцшу и др. [126] на молодняке крупного рогатого окота и овцах, а также Леделя (1970) на молодняке крупного рогатого скота оказалось, что бедный фосфором опытный рацион хотя и приводил к типичным симптомам фосфорной недостаточности, таким, как изменение костей и снижение уровня фосфора в плазме крови, однако в отношении плодовитости различий между группами не было. Как показывает Гегель на табличном материале, между показателями плодовитости двух опытных и одной контрольной групп молодняка крупного рогатого скота достоверных различий не было. Телята от животных, содержавшихся на дефицитных по фосфору рационах, имели ту же массу при рождении, что и телята двух других опытных групп (табл. 13).
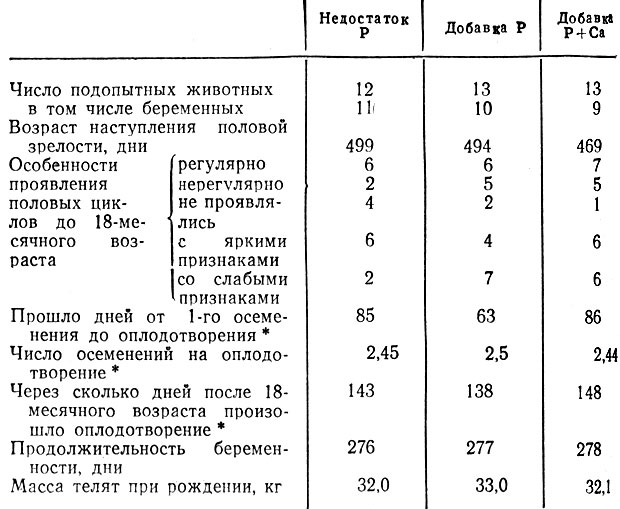
Таблица 13. Показатели плодовитости телок при разном уровне обеспеченности фосфором и калием
* (Среднее по беременным животным.)
Результаты исследований на телках подтвердились в опытах на овцах. Хотя уменьшение количества карма на 30% привело к резкому спаду плодовитости, разница в уровне обеспеченности фосфором не проявилась. Беременными становились те животные, которые позднее погибли из-за недостатка фосфора или обнаруживали клинически заметные изменения костей. После содержания в течение 11/2 лет на рационах, дефицитных по фосфору, которое привело к заметному ухудшению развития и роста, ритм половых циклов не изменился [127]. Исследование матки и яичников, проведенное по окончании опытов, не дало указаний на неблагоприятное влияние недостатка фосфора на функции женских половых органов [125]. С другой стороны, эти авторы [127] предполагают непрямую связь между уровнем обеспеченности фосфором и нарушением воспроизводства.
Эти и последующие исследования отчетливо показали, что фосфор отнюдь не играет доминирующей роли в процессах воспроизводства крупного рогатого скота, важен общий минеральный статус организма. По меньшей мере три десятка лет ученые чрезмерно ограничивались анализом этого элемента, не учитывая действия других макро- и микроэлементов. В настоящее время, по мнению ученых, работающих в сотрудничестве с Аике и Хольцшу, мы не имеем никаких четких указаний на то, что несбалансированность кальция и фосфора и гипофосфатозы специфически воздействуют на половую систему. Наблюдаемые при недостатке фосфора нарушения плодовитости надо рассматривать как следствие общего нарушения обмена веществ, в особенности недостатка энергии, как выработанный, очевидно, нейроэндокринной системой синдром приспособления к плохим условиям кормления. Так, при дефиците фосфора в значительной степени снижается сопротивляемость организма инфекционным заболеваниям, ухудшаются упитанность, поедаемость корма и способность к превращению каротина в витамин А, что может обусловить бесплодие. Отсюда должно быть ясно, что любая дальнейшая несбалансированность в питательных веществах усиливает это действие и что чувствительность к нефтизиологическому обеспечению кальцием и фосфором подвержена большим индивидуальным и виутрипородным колебаниям.
Оптимальным для крупного рогатого окота надо считать соотношение Са : Р от 1 : 1 до 2 : 1 (1 СаО : 1 Р205 ≈ 1,6 Са : 1 Р). В обычных для ГДР условиях кормления животных изменение этого соотношения до 3 : 1 и даже 4 : 1 не может принести вреда, если в рационе достаточно фосфора или витамина D. В противоположном направлении сходятся литературные данные об отношении 0,3 : 1. Вначале при расширении соотношения между кальцием и фосфором с 1 : 1 до 3 : 1 отмечалось даже улучшение показателей воспроизводства (рис. 10).
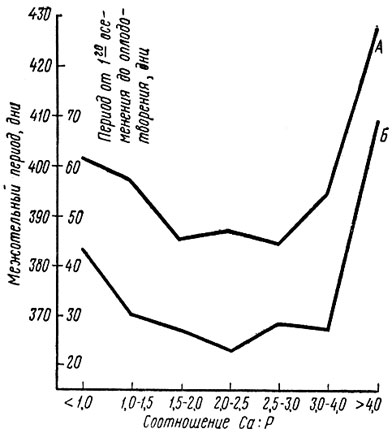
Рис. 10. Продолжительность межотельного периода (А) и интервал от первого осеменения до оплодотворения (Б) у коров при различном соотношении Са : Р в рационе (по Конерманну)
При дальнейшем расширении за пределы 3 : 1 произошло удлинение межотельного периода и интервала от первого осеменения до оплодотворения. Самыми худшими оказались результаты при соотношении Са : Р сверх 4 : 1 [149]. Однако сужение этого соотношения обеспечивается не одним лишь высоким уровнем фосфора в почве, хотя как раз от него зависит состав травостоя на лугопастбищных угодьях и тем самым минеральный статус животных. В самой почве тоже должны быть созданы предпосылки для того, чтобы в процессе промежуточного обмена фосфора при участии азотфиксирующих бактерий происходило его накопление в кормовых растениях. Этот процесс в значительной степени зависит от рН почвы. При оценке ситуации с воспроизводством, как указывают Хельферих [100] и Конерманн [149], необходимо исследовать не только абсолютное содержание фосфора и кальция и их соотношение между собой, но и долю этих элементов в рационе в расчете на 100 г переваримого протеина, так как именно между плодовитостью и долей фосфора на 100 г переваримого протеина должна существовать тесная корреляция. Самые лучшие показатели воспроизводства Конерманн установил при содержании 3,5-3,9 г фосфора на 100 г переваримого сырого протеина. Отклонения в сторону как повышения, так и понижения этой величины имели следствием значительное удлинение межотельного периода и интервала от первого осеменения до оплодотворения (рис. 11). То же наблюдалось и у животных, получавших недостаточно фосфора. Разумеется, в этом случае доля фосфора в расчете на 100 г переваримого сырого протеина была ниже оптимума - всего 3-3,5 г. Отсюда ясно, что соотношение питательных веществ должно оцениваться не изолированно, а в связи с абсолютными цифрами потребления питательных веществ.
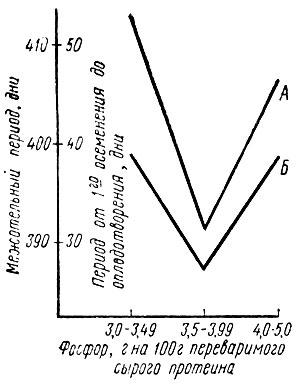
Рис. 11. Продолжительность межотельного периода (А) и интервал от первого осеменения до оплодотворения (Б) в зависимости от соотношения фосфор : переваримый сырой протеин в рационе при избытке фосфора более 10 г на животное в день (по Конерманну)
Поэтому добавка 10 г фосфора в рацион животных при дефиците этого элемента вызывала сокращение интервала между первым осеменением и оплодотворением на 10 дней, если соотношение между фосфором и переваримым сырым протеином оставалось 3,5 : 100. Такие же добавки фосфора, но из расчета 4,5 на 100 г переваримого сырого протеина, вызывали удлинение этого интервала до 26 дней. Следовательно, благотворное само по себе действие добавок фосфора при его недостаточности может быть полностью снято или даже превратиться в свою противоположность. Очевидно, что характер действия целенаправленной подкормки определяется также сложившейся в каждом конкретном случае кормовой ситуацией. Может быть именно этим объясняются в известной степени результаты, полученные Легелем (1970) и представленные им в таблице 13, из которой видно, что у животных с недостатком фосфора в рационе (отношение фосфора к переваримому протеину = 2,0/100 г) показатели плодовитости были не намного хуже, чем у животных, получавших рацион с добавкой фосфора до уровня потребности, но все еще не с оптимальным отношением фосфора к переваримому протеину - 3,2 г на 100 г.
Отношение кальций: сырой протеин считается оптимальным, если на 100 г переваримого сырого протеина приходится 7-8 г кальция [149].
Более высокие дозы кальция неблагоприятно сказывались и на плодовитости (рис. 12). При пересчете на условия опытов Легеля (1970) это отношение для группы с недостатком фосфора должно составить 9,7/100 г, для группы, получавшей добавку фосфора,- 8,7/100 г и для группы, получавшей добавку кальция и фосфора,- 16,4/100 г.
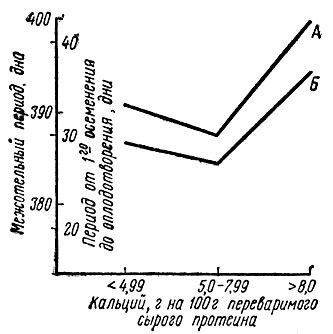
Рис. 12. Продолжительность межотельного периода (А) и интервал от первого осеменения до оплодотворения (Б) при различном соотношении Са : переваримый сырой протеин в рационе коров (по Конерманну)
Относительно обеспечения сельскохозяйственных животных фос4 ором за счет кормов можно сказать, что такие кормя, как пастбищная трава, зеленый корм, сено, силос и травяная мука и другие корма собственного производства, доля которых в рационах все еще значительна, даже при регулярном удобрении почвы фосфатами большей частью бедны фосфором и относительно бедны кальцием. Особенно это относится ко всем растениям в период интенсивного роста. В дальнейшем такое положение осложняется тем, что из хозяйственных кормов усваивается максимум 20% содержащейся в них извести и 50% фосфора [13]. Отложения лигнина, которые увеличиваются по мере созревания растений, к концу пастбищного сезона уменьшают доступность фосфора и других содержащихся в кормах питательных веществ. Вопрос о том, как влияет погода во время роста растений на содержание в них фосфора и кальция, все еще полностью не выяснен. Результаты опытов Неринга [192], показавшие, что при сухой погоде содержание фосфора в растениях уменьшается, а кальция возрастает (при влажной и холодной наоборот), не подтвердились исследованиями Ондершека [196], который установил снижение содержания минеральных веществ в растениях в выраженные периоды засухи, как этого и следовало ожидать вследствие низкой растворимости в воде почвенных солей. Но в кормах, полученных в самые богатые осадками периоды, содержание минеральных веществ оказалось самым низким [256].
Приводимые в литературе данные о нормах потребности в фосфоре сельскохозяйственных животных нередко довольно значительно различаются, так как эта потребность может быть выражена как в доступном, так и в общем фосфоре. Нередко указывают потребность в фосфоре, относя ее к разным исходным величинам, например на животное в день, в процентах сухого вещества корма, на живую массу или продуктивность животного. Данные в граммах на животное в день, как и в процентах сухого вещества корма, одинаково непригодны, если при этом не указаны масса и продуктивность животных. Поэтому неудивительно, что, например, для крупного рогатого скота были выведены формулы, с помощью которых определяется конкретная потребность. По Ван дер Мейлену и Френсу (1959), потребность в обоих элементах для высокопродуктивных коров определяется по следующей формуле:
где ГУ - годовой удой в тыс. кг и СУ - суточный удой в кг. Таким образом, для коровы с годовым удоем 4000 кг и суточным 15 кг потребность в кальции и фосфоре составит:
Для вычисления потребности в фосфоре молодняка крупного рогатого скота в период активного роста Хольцшу и Мартин [128] вывели следующее отношение:
Однако при широком отношении Са : Р эту норму следует увеличить на 15%.
Как и многие другие авторы, Шмидт [220], Хольцшу и др. [129] тоже указывают, что изменение уровня обеспечения животных фосфатом очень быстро сказывается на содержании неорганического фосфора в сыворотке крови. Поэтому исследованием крови удобно воспользоваться для оценки уровня обеспеченности фосфором стада в данный конкретный момент. Для суждения же об этом показателе в рамках какого-либо определенного длительного промежутка времени такие исследования необходимо проводить регулярно. Уровень фосфора в сыворотке крови не всегда соответствует абсолютному поступлению его с кормом, но он достаточно хорошо отражает степень удовлетворения потребности в нем. Добавки кальция на уровень фосфора не влияют и не оказывают также самостоятельного влияния на уровень кальция. Дальнейшие взаимоотношения между кальцием и неорганическим фосфором в сыворотке крови можно объяснить главным образом недостатком фосфора, а не просто слишком широким отношением Са : Р, как это нередко предполагают. Наряду с исследованиями сыворотки крови в качестве индикатора обеспеченности организма фосфором могут служить анализы волосяного покрова и костей.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://animalialib.ru/ 'Животноводство'