Секреция молока
Наступление секреции молока, или лактогенеза, протекает под влиянием гормонов аденогипофиза (особенно пролактина и соматотропина), которые непосредственно влияют на альвеолярный эпителий, при одновременном участии кортикоидов и тироксина. Обильная секреция наступает в связи с родами вследствие снижения уровня прогестерона (из плаценты) по отношению к уровню эстрогена в организме. После родов секрецию молока акти-визирует аденогипофиз. Однако наступление секреции трудно объяснить только эндокринными факторами. Существенна роль нервной регуляции, вероятно, вследствие импульсов из шейки матки, а также регулярных импульсов из молочной железы, особенно во время сосания и доения.
Анализируя регуляцию установившейся лактации (лактопоэза), необходимо рассматривать три группы процессов: регуляцию секреторной деятельности молочной железы, поддержание ее структуры и взаимодействие функции железы с другими системами целого организма.
Последующая интенсификация и поддержание молокообразования в сформированной железе связаны с выделением гормонов передней доли гипофиза (пролактина, соматотропина), которое регулируется нервной системой, а также с участием гормонов коры надпочечников, щитовидной и половых желез. Выделение пролактина активизирует акт сосания и доения коров, повидимому, через нейрогормональные стимулы. В ходе лактации содержание пролактина в гипофизе снижается параллельно снижению секреции молока. При этом в начале лактации возбуждение столь сильно, что организм коровы для выработки молока часто вынужден использовать резервы собственных тканей.
Схематически нейрогормональную регуляцию лактации коров можно представить так:
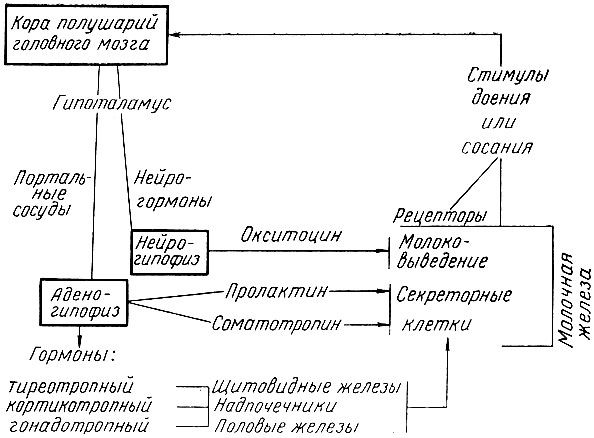
В серии опытов по денервации молочной железы выявлено заметное снижение удоев. Этим подтвердилось участие нервной системы в регуляции секреции молока. Центральная нервная система может влиять на молокообразование через гипоталамо-гипофизарную систему путем изменения обмена веществ в организме и в молочной железе. Роль центральной нервной системы в регуляции лактации отчетливо проявляется в зависимости молочной продуктивности от типологических особенностей высшей нервной деятельности. Э. П. Кокорина показала, что у животных сильного уравновешенного подвижного типа по сравнению с коровами слабого, неуравновешенного типа выше молочная продуктивность и устойчивость лактационной кривой, меньше суточные колебания удоев.
Гистологическими и биохимическими исследованиями выявлено, что железистые клетки забирают из крови определенные вещества для синтеза составных частей молока. Изучением различий в составе артериальной и венозной крови вымени выяснены основные предшественники молока.
Витамины, минеральные вещества, энзимы из прилегающих кровеносных капилляров избирательно проникают через мембрану альвеол в их полости и становятся составными частями молока.
Биосинтез белков молока происходит из совокупности свободных аминокислот плазмы, а также из белков и полипептидов притекающей крови.
Синтез а и р-казеина и р-лактоглобулина (составляют 90% белков коровьего молока) происходит в молочной железе.
Иммуноглобулин и молочный сывороточный альбумин поступают в молоко, возможно, из крови, Глобулин молозива может попадать в него из клеток плазмы молочной железы.
Синтез лактозы происходит из свободной глюкозы и глюкозы крови. Содержание глюкозы в молоке относительно более постоянное, чем содержание других компонентов.
Биосинтез жиров молока осуществляется из глицерина, жирных кислот, нейтрального жира и фосфолипидов крови.
Образующаяся в преджелудках при микробном расщеплении углеводов уксусная кислота в молочной железе может превращаться в глицерин и жирные кислоты.
Поэтому изменения интенсивности брожения в рубце при рационах разного состава (по сахаро-протеиновому отношению и количеству переваримой клетчатки) вызывают заметные колебания процента жира в молоке. Жирномолочные коровы отличаются более интенсивным брожением и накоплением летучих жирных кислот в рубце, чем жидкомолочные. Активизация щитовидной железы во время зимних холодов повышает жирообразование.
Торможение активности этой железы во время летней жары, а также при скармливании коровам в большом количестве капусты и турнепса, содержащих антитиреоидные вещества, снижает жирность молока. Выведение жира из клеток железистого эпителия заметно тормозится при значительном повышении внутривыменного давления через 8-10 часов после дойки. Поэтому при коротких интервалах между дойками содержание жира в молоке выше, чем при больших, например, после ночного перерыва.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://animalialib.ru/ 'Животноводство'