Молоковыведение и внутривыменное давление
Молоковыведение, или молокоотдача в физиологическом понимании - согласованные рефлекторные реакции миоэпителия и гладкой мускулатуры вымени, стенок и сфинктера соска в ответ на соответствующие раздражения кожи сосков и вымени в определенной обстановке (при доении или сосании теленком). Молокоотдачу характеризуют скрытый период рефлекса, изменение внутривыменного давления и скорости молоковыведения, а также различная степень торможения, или "задержки", молока. Молокоотдача протекает синхронно во всех четвертях вымени. Нервная система оказывает значительное влияние на эти процессы.
Рефлекс молокоотдачи протекает в две фазы. Первая фаза имеет небольшой латентный период (1-4 сек.) и характеризуется расслаблением гладкой мускулатуры цистерны и протоков, кратковременным понижением внутрицистернального давления (происходит подготовка к пропусканию и принятию молока из альвеол). Рефлекторная дуга в первой фазе начинается с рецепторов соска (рис. 7) и по афферентным волокнам задних корешков (1) проходит к спинному мозгу, далее через дорсальные и боковые канатики (2) спинного мозга к медиальным петлям (3) продолговатого мозга. По стволу мозга импульсы достигают супраоптических ядер (4) гипоталамуса, затем коры больших полушарий (5). От супраоптических ядер начинаются эффективные волокна, которые в составе супраоптико-гипофизарного 6) достигают нейрогипофиза (7). Нейрогипофиз в ответ на сосание или доение выделяет в кровь гормон окситоцин, который вызывает сокращение миоэпителия альвеол (8). Имеются данные, что миоэпителий (9) может сокращаться в ответ на механические раздражения железы. На уровне поясничных сегментов спинного мозга происходит переключение импульсов с афферентного на эфферентный путь, представленный симпатическими нервами молочной железы.
Вторая фаза - нейрогормональная - более длительна и связана с действием окситоцина на миоэпителий альвеол. Латентный период второй фазы около 35-40 сек.
Под влиянием окситоцина (или комплекса гормонов - питуитрина) резко сокращается миоэпителий альвеол, изгоняя молоко в протоки. При этом протоки сокращаются, отчего увеличивается их просвет, облегчая переход молока в цистерну.
Сокращение мускулатуры крупных протоков, цистерны и сфинктера соска регулируется эффективными эфферентными нервами молочной железы, а не окситоцином. Эфферентные пути наружного семенного нерва состоят из нервных волокон, вызывающих спазмы и сокращение гладкомышечных волокон, и из волокон, вы-зывающих расслабление гладкой мускулатуры.
Благодаря быстрой выработке условных рефлексов на обстановку (стереотип) доения молоко из альвеол и мелких протоков к моменту доения переходит в цистерну, из которой быстро выдаивается.
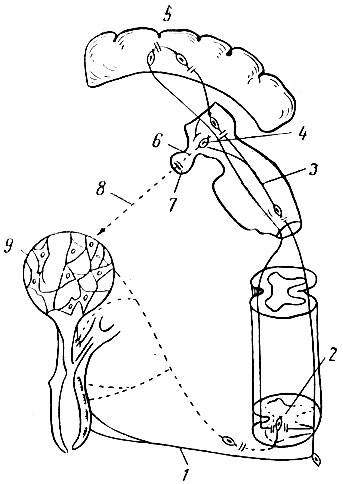
Рис. 7. Схема нейрогормонального рефлекса молокоотдачи (по М. Г. Заксу, 1958): 1 - афферентные волокна задних корешков; 2 - дорсальные и боковые канатики спинного мозга; 3 - медиальные петли продолговатого мозга; 4 - супраоптические ядра гипоталамуса; 5 - кора больших полушарий; 6 - супраоптикогипофизарный тракт; 7 - нейрогипофиз; 8-действие окситоцина, выделяемого нейрогипофизом; 9-миоэпителий альвеол
Сложный условцорефлекторный процесс молокоотдачи связан с повышением активности всех частей вымени, его кровеносных и лимфатических сосудов, а также многих функций организма. Усиливается приток крови и повышается температура кожи вымени.
Э. П. Кокорина установила четкую связь характера молокоотдачи с типом нервной деятельности. У коров с высокой подвижностью нервных процессов скорость выдаивания более постоянна, чем у коров с плохой подвижностью нервных процессов. По молокоотдаче можно судить о силе, уравновешенности и подвижности нервных процессов.
Торможение молоковыведения осуществляется рефлекторным сокращением гладкой мускулатуры устьев протоков, а также нейрогормонально путем выделения адреналина надпочечниками. Адреналин сужает молочные протоки и тормозит освобождение окситоцина.
Для практики доения наиболее важно условнорефлекторное торможение молокоотдачи, возникающее при нарушении стереотипа доения: грубое обращение с коровами, испуг и болевые раздражения сосков, возбужденность во время охоты, перемена места, очередности и времени выдаивания, смена доярки, наличие шума, посторонних лиц и др. Это торможение может быть сильным, средним или слабым. Более сильное торможение чаще проявляется у животных слабого типа нервной деятельности (вплоть до торможения секреции).
Между секрецией, молоковыделением и молоковыведением существует тесная взаимосвязь и согласованность. Эффективность молоковыведения зависит от степени заполнения вымени и соответственно от распределения молока в отделах емкостной системы и достигнутого внутрицистернального давления. Эти условия и регулярные опорожнения вымени влияют на скорость и харак: тер секреции.
По данным Ю. М. Огороднего, стимуляция молокоотдачи одновременно вызывает интенсивное выведение жира и других веществ альвеолярным эпителием, что ведет к быстрому и значительному повышению жирности молока последующих порций удоя. Как полагает Г. И. Азимов, это явление происходит с некоторым запозданием- к концу дойки и после нее.
Горячие обмывания и массаж вымени стимулируют нейрогормональное открытие протоков и более полное жировыведение при доении (Ю. М. Оленов и Н. А. Ниукканен). Быстрое выдаивание обеспечивает более полное опорожнение альвеолярной зоны вымени и обогащение молока жиром как за дойку, так и за всю лактацию. Отдельным коровам присуще свойство увеличивать жировыведение при энергичной молокоотдаче. Это способствует одновременному повышению удоев и жирности молока. Такое свойство представляет интерес для селекции. Всякое торможение молокоотдачи ведет к снижению продуктивности коров и сокращению у них лактационного периода (самозапуску).
Рефлекс молоковыведения можно характеризовать по латентному периоду, соотношению порций удоя и остаточного молока, изменению цистернального давления и продолжительности или скорости (интенсивности) молокоотдачи.
Латентный период - время от начала стимуляции до активного изгнания секрета из железы в виде непрерывной струи молока - подвержен значительным индивидуальным колебаниям и длится от 15 до 120 сек. и больше.
Длительность скрытого периода весьма изменчива. Она неодинакова у разных животных и может меняться у одного и того же животного, в ходе лактации постепенно увеличиваясь; зависит от обстановки, или стереотипа доения, степени наполнения вымени молоком (интервалов дойки), функционального состояния железы и возбуждения нервной системы коров.
В опытах А. А. Сюсюкина выяснилось, что чем выше разовый удой в различные дойки, тем жороче скрытый период рефлекса молоковыведения. В начале и в разгаре лактации коров рефлекс молоковыведения протекал бурно и латентньш период в среднем был короче (48- 49 сек.), чем к концу лактации (86-96 сек.). Поэтому во второй половине лактации необходимо более интенсивное и продолжительное стимулирование, чтобы добиться энергичной молокоотдачи и полного опорожнения вымени при одновременном сокращении времени доения коров.
Соотношение пропорций удоя определяют с помощью катетеризации сосков. Сначала без стимуляции молоковыведения пассивно вытекает через катетер цистернальная порция. Затем производят массаж и доение остальных сосков. В результате сокращения альвеол и мелких протоков через катетер получают альвеолярную, или рефлекторную, часть удоя. После инъекции окситоцина доением извлекается остаточное, или резидуальное, молоко.
Многочисленными исследованиями установлено, что альвеолярная порция относительно устойчива, тогда как цистернальная часть удоя значительно варьирует. Их соотношение изменяется в зависимости от индивидуальных различий животных и стадий лактации, возраста коров, интервала и стереотипа доек, емкости вымени, цистернального давления и др. Это затрудняет (наряду с технической сложностью) определение и интерпретацию получаемых данных в условиях ферм. Обычно такие опыты проводятся при ручном выдаивании небольшого числа коров, что ограничивает использование полученных данных в селекционной работе.
Изучая взаимосвязи порций удоя при двух и трехкратной дойке коров, В. В. Кулик установил, что объем альвеолярной порции относительно устойчив в течение ряда месяцев лактации, а доля цистернальной порции в удое колебалась и зависела от интервалов дойки (табл. 3).

Таблица 3. Объем цистернальной порции в зависимости от интервалов
П. В. Полетаев, исследуя особенности рефлекса молоковыведения у высокоудойных, но с разной жирностью молока коров холмогорской и ярославской пород на 2-5-м месяце лактации, нашел, что в утреннюю дойку у жирномолочных коров объем цистернальной порции в 3,25 раза больше альвеолярной, а у жидкомолочных эти объемы почти одинаковы. Наоборот, альвеолярная порция у жирномолочных коров в 2-2,5 раза меньше, чем у жидкомолочных. Оказалось также, что чем выше удой, тем больше процент цистернальной порции.
Наши исследования внутривыменного давления перед дойкой и его изменений под влиянием стимуляции молокоотдачи, а также литературные данные по этому вопросу позволяют сделать следующие обобщения. Давление пропорционально количеству молока в железе и отражает степень ее заполнения. В то же время оно имеет значительные колебания у одних и тех же коров от дойки к дойке.
Под влиянием стимуляции внутривыменное давление заметно повышается. Рефлекторное сокращение миоэпителия альвеол обеспечивает быстрое изгнание секрета в цистерну железы, расширяет сосковую цистерну и облегчает раскрытие соокового канала, возможно, через рефлекторное расслабление сфинктера. Однако сила отрицательного давления под соском примерно в 10 раз превышает положительное давление в цистерне соска (380:36) в начале дойки и еще больше в конце. Поэтому основное значение внутривыменного давления и его повышения после стимуляции молокоотдачи сводится к быстрому заполнению цистерны железы и соска молоком, которое затем извлекается при сосании или доении. Для более полного опорожнения альвеолярной зоны вымени весьма существенно, чтобы скорость доения соответствовала интенсивности молокоотдачи.
Внутрицистернальное давление в начале дойки уменьшается медленно, а к концу ее быстро (рис. 8), соответственно снижается и скорость молокоотдачи. Однако снижение давления в цистерне отстает от темпа опорожнения вымени. Благодаря этому поддерживается интенсивная молокоотдача. Перекрест линий давления и процента выдоенного молока у первотелок отмечен раньше при 70%, у коров второй лактации позже - при 60% выдоенности, что обусловлено различиями их удоев.
По мере выдаивания и снижения давления спадается кожа цистерны и сосков вымени. Наличие невыдоенного молока в отдельных четвертях легко установить прощупыванием цистерны.
Сосковый канал и сфинктер оказывают сильное влияние на скорость молокоотдачи. Растяжимость канала-соска зависит от строения кончика соска, объема и соотношения гладких мускулов и эластичных волокон соединительной ткани, а также от тонуса сфинктера. Перечисленные свойства наследственные и в разных сочетаниях обусловливают степень тугодойности или легкодойности коров. Диаметр соскового канала с возрастом коров увеличивается, отчего повышается и скорость молокоотдачи.
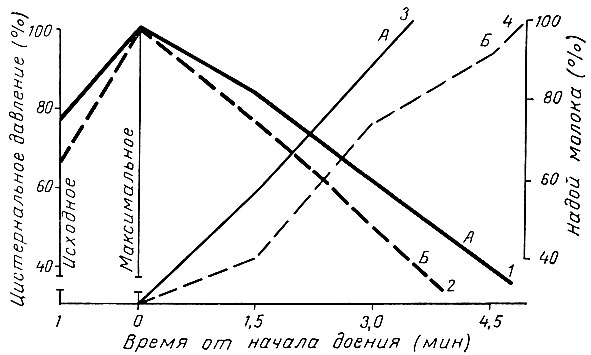
Рис. 8. Внутрицистернальное давление в процессе машинного доения: А - первая лактация; Б - вторая лактация; 1, 2 - давление; 3, 4- выдоено молока
У отдельных коров со слабым сфинктером еще до начала дойки наблюдается непроизвольное истечение молока. Это существенный дефект, при котором происходит потеря продукции.
У тугодойных коров замедленно выделяется альвеолярная порция молока, отчего нередко увеличивается объем резидуальной порции, сокращается продолжительность лактации и снижаются удои. На выдаивание таких коров требуются большие затраты труда и времени.
Резидуальное, или остаточное, молоко, извлекаемое по окончании нормальной дойки после введения 18-20 не окситоцина, регулярно депонируется и является причиной колебаний удоя и жирности молока по дойкам за день, за смежные сутки и за всю лактацию коров. Количество резидуального молока и жира -наследственный признак и у разных коров составляет от 5 до 25% и более общего удоя. Объем остаточного молока изменяется в ходе лактации и от продолжительности интервалов между дойками, и его количество пропорционально удою. Увеличение количества остаточного молока характеризует торможение молокоотдачи или полноту выдаивания, снижает продуктивность коров. Массовые определения остаточного молока пока затруднительны и в селекции не используются. Однако при исследованиях емкости вымени и колебаний скорости молокоотдачи, скорости секреции и состава молока при различных интервалах доек определять его необходимо.
На практике нерегулярное и неполное выдаивание, а также замедленное опорожнение вымени при доении ведет к снижению удоя и жирности молока, более крутому падению лактационной кривой и сокращению числа дней лактации, к преждевременному запуску коров. Различными приемами стимуляции молокоотдачи можно повышать полноту выдаивания, удои и жирность молока, хотя и в разной степени у отдельных коров.
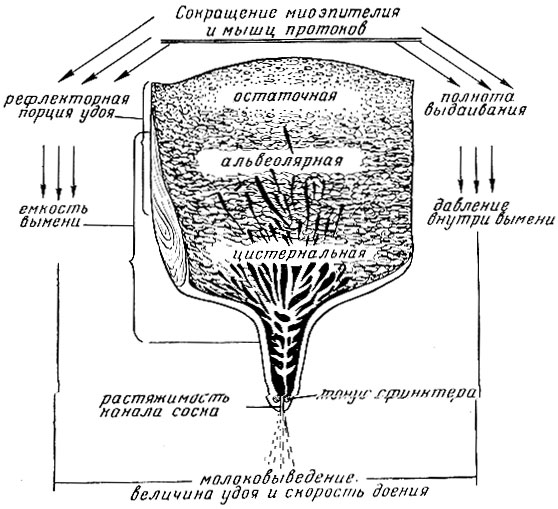
Рис. 9 Характеристики процессов молокоотдачи: сокращения миоэпителия и гладких мышц протоков обусловливает рефлекторную порцию удоя и полноту выдаивания, которые вместе отражают емкость вымени. Скорость доения зависит от величины удоя, давления внутри вымени, растяжимости канала соска и тонуса сфинктера
Изложенные характеристики молокоотдачи (рис. 9) и их взаимосвязи (табл. 4) необходимо знать и исполь-зовать при организации доения и селекционной оценке коров по их пригодности для двухразовой машинной дойки. Кроме того, нужно знать взаимосвязи молочной железы в целостном организме.

Таблица 4. Влияние компонентов рефлекса молоковыделения на суммарные его показатели - скорость доения и удой коров
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://animalialib.ru/ 'Животноводство'